Abstract
Epigenetic inheritance alerts naïve descendants to prepare for stresses that could still be present, whereas distant descendants return to a basal state after several generations without stress. However, organisms are frequently exposed to stresses successively across generations. We found that parental hypoxia exposure increased P0 longevity, caused intergenerational lipid reduction, and elicited transgenerational fertility reduction that was depen dent on generationally transmitted small RNAs. Here, we find that Caenorhabditis elegans adapt to repeated generational stresses. We show that, upon two repeated generational hypoxia exposures, the life- span extension is eliminated, and after four repeated generational hypoxia exposures, the reduced fertility is eliminated. Trans generational adaptation also occurred in response to changes in glucose availability. Transgenerational hypoxia adaptation is dependent on the H3K27 trimethyltransferase PRC2 complex, and we identified transgenerationally adapted genes. Our findings reveal that transgenerational adaptation occurs and suggest that H3K27me3 is a critical modification for adapting to repeated generational stresses.
INTRODUCTION
Most genetic information is encoded and inherited through DNA (1). However, previous findings demonstrated that changes in gene expression can be mediated in the absence of changes to the DNA sequence. Heritable phenotypes regulated by epigenetic information include physical appearance (2, 3), metabolism (4), behavioral state (5), and longevity (6, 7). The phenomena of heritable nongenetic in formation being transmitted across generations presumably evolved to allow organisms to pass important information to their descen dants, allowing their progeny to better survive altered environments without mutations to their genome. By avoiding mutation, the or ganism can survive irregular conditions but returns to a basal state when the environmental conditions improve (8). However, how does the adaptable nature of epigenetics (9) respond to repeated en vironmental stresses? Can repeated environmental stresses override the canonical epigenetic inheritance? Is there adaptation on a trans generational scale as well? Although laboratory experiments have mostly focused on con ditions of a transient environmental stress followed by a return to basal environmental conditions, nature rarely follows such con trolled conditions. Organisms are frequently exposed to stresses repeatedly over their lifetime or over a generational timescale or al ternatively exposed to one stress in one generation and then a sec ond stress in a subsequent generation. Here, we asked whether repeated generational exposure to stresses altered the generational inheritance clock, overrides canonical epigenetic inheritance, or changed the magnitude of the phenotypic response.
Low oxygen (hypoxia) is an environmental stress, which can be harmful in many contexts such as inducing ischemia and eventually cell death (10). The hypoxic tumor microenvironment is also correlated with increased metastasis and mortality (11). Fetal hypoxia impairs de velopment and is associated with several pregnancy disorders and complications (12–15). Ocean warming and acidification caused by climate change have lowered ocean oxygen levels and therefore are threatening marine life (16–18). In mammalian cells, in response to hy poxia, the H3K27me3 demethylase KDM6A/UTX senses lowered oxy gen levels, causing elevated H3K27me3 levels and blocks cellular differentiation (19). Caenorhabditis elegans frequently encounter oxy gen depletion in nature when the soil gets flooded (20). We and others have previously found that hypoxia increases C. elegans life span, re duces neutral lipids, and reduces fertility in the exposed generation (P0) (21–24). We previously found that hypoxia exposure in the P0 generation causes an intergenerational (F1) reduction in neutral lipids and a transgenerational (F1+F2) reduction in fertility accompanied by a dysregulation of gene expression (24). We found that the transgenera tional epigenetic inheritance of hypoxia- induced fertility defects was dependent on the small RNA argonaute HRDE- 1, the putative H3K9 trimethyltransferase SET- 25, the putative H3K27 trimethyltransferase MES- 2, and the putative H3K27me3 demethylase JMJD- 3.2 (24). We further found that small RNAs isolated from hypoxia- treated C. elegans are sufficient to elicit fertility defects and identified transgenerationally dysregulated small RNAs. We identified that one specific double- stranded RNA (dsRNA) against F44E5.4/5 is sufficient to induce a heritable decrease in fertility and is directly transmitted from parents to their progeny (24). However, it is still unclear whether animals will adapt to repeated generational exposure to hypoxia.
Here, we show that C. elegans that were exposed to hypoxia re peatedly over several generations failed to display hypoxia- induced phenotypes. We found that upon two repeated generational exposures to hypoxia, C. elegans no longer displays an increase in life span, and after four repeated generational exposures to hypoxia, C. elegans no longer displays a decrease in reproduction. In addition, this transgen erational adaptation was not unique to hypoxia—exposure to 1% glucose caused a subtle increase in neutral lipids and a decrease in fertility, whereas two repeated generational exposures to elevated glucose eliminated the capacity of C. elegans to respond to the ele vated glucose to alter lipid content; after three repeated generational exposures to glucose, C. elegans no longer displays a decrease in reproduction. The transgenerational adaptation of decreased reproduction to hypoxia is dependent on the putative H3K27 trimethy transferase PRC2 complex, and we identified critical genes that adapted on a transgenerational time frame to repeated hypoxia expo sure. Together, our results reveal that transgenerational adaptation oc curs and suggest that H3K27me3 is a critical modification for adapting transgenerationally to repeated generational stresses.
RESULTS
C. elegans adapt to repeated hypoxia exposure
Hypoxia exposure increases life span and decreases fertility (24, 25). To determine whether C. elegans can transgenerationally adapt to stresses, we repeatedly exposed C. elegans to hypoxia across multiple generations (Fig. 1A) and measured reproduction and longevity. We exposed larval stage (L4) worms to hypoxia (0.1% oxygen) for 16 hours before returning to a normoxic environment (~21% oxygen). We subsequently exposed F1, F2, F3, and F4 descendants to similar hypoxia periods (Fig. 1A) and measured progeny production at each generation. We found that the P0, F1, and F2 generations all dis played reduced fertility in response to hypoxia treatments; however, by the fourth repeated generational exposure to hypoxia, C. elegans no longer displayed a hypoxia- induced fertility decrease (Fig. 1B and table S1). This adaptation persisted as the fifth repeated generational exposure to hypoxia also no longer displayed a hypoxia- induced fer tility decrease. We found that the generational adaptation to hypoxia A B Generation occurred more rapidly for longevity. Whereas the initial exposure to hypoxia subtly increased C. elegans life span (Fig. 1C and table S2; 13% P = 0.0022 by combined Fisher’s test), as has been shown previ ously (24, 25), C. elegans exposed to hypoxia, whose parents had also been exposed to hypoxia, did not display an increase in life span (Fig. 1D and table S2; P = 0.1495 by combined Fisher’s test). To gether, these findings suggests that C. elegans transgenerationally adapt to become resistant to hypoxia conditions after repeated gen erational exposure.
C. elegans adapt to repeated changes in glucose availability
To determine whether transgenerational adaptation is specific to hy poxia or is a general phenomenon, we exposed repeated generations of C. elegans to either 1 or 2% glucose. A 2% glucose exposure had previously been demonstrated to reduce the life span of the exposed generation (26–29) and cause a transgenerational reduction in fertility for three generations (29) and an intergenerational increased resis tance to oxidative stress and protection from neurodegeneration (29). We found that C. elegans exposure to elevated glucose led to an in crease in neutral lipids as assessed by oil red O staining (Fig. 2, A and B, and table S3) in the exposed generation. This parental exposure induced a decrease in neutral lipid content in the F1 generation (Fig. 2, C and D, and table S3), suggesting that elevated parental glucose con tent causes intergenerational decreased neutral lipids. Similar to what we had found with the effect of repeated hypoxia exposure, we found that repeated generational exposure to elevated glucose eliminated the
second generation’s capacity to respond to elevated glucose (Fig. 2, E and F, and table S3). In addition, we found that the decrease in fertility observed in response to elevated glucose (Fig. 2G and table S4) was still present after a successive generational exposure to elevated glu cose (Fig. 2H and table S4) but was eliminated in the third repeated generational exposure to elevated glucose (Fig. 2I and table S4). To gether, these results suggest that C. elegans adapt with regard to their fat content and fertility in response to repeated exposures to elevated glucose similar to what we had observed with life span and fertility in response to repeated hypoxia exposure (Fig. 1). These findings suggest that transgenerational adaptation is a general phenomenon in response to repeated exposure to environmental stresses.
Transgenerational adaptation to hypoxia requires genes involved in metabolism, immunity, and the stress response
To determine how C. elegans adapted to repeated generational exposures to hypoxia, we performed RNA sequencing (RNA-seq) of normoxia- reared controls and C. elegans exposed to hypoxia for one, two, three, or four generations by which point all hypoxia- induced phenotypes we tested were no longer responsive to hy poxia treatment. We found that hypoxia treatment induced similar gene expression changes as we had previously observed (fig. S1A; r2 = 0.96) (24), suggesting that our RNA-seq analysis and hypoxia treatment were consistent and reproducible. We found that hypoxia treatment in a single generation caused an up- regulation of 2222 genes and a down- regulation of 1483 genes, which reached our sig nificance cutoff across multiple datasets (Fig. 3A). These genes were enriched for genes involved in a variety of metabolic processes, im mune response, and lipid catabolism (fig. S2 and table S5). We found that hypoxia treatment elicited mostly similar changes in gene ex pression if performed for one, two, three, or four successive gen erations (Fig. 3, A and B, and fig. S1B). We were most interested in genes whose altered expression disappeared when the transgenera tional phenotypes displayed adaptation, in the second generation for longevity and the fourth generation for fertility. These genes would include those that were dysregulated in response to hypoxia
(42%) A Replicate: 123123 Normoxia Hypoxia Generation of exposure: E 1 Stress response to metal ion Stress response to copper ion Response to copper ion Response to metal ion Fatty acid -oxidation Apoptotic signaling pathway Peptide metabolic process Mitochondrial protein processing Peptide biosynthetic process Fatty acid oxidation Lipid oxidation Lipid transport Stress response to cadmium ion Mitochondrial translation Amide biosynthetic process Sterol transport Response to inorganic substance 0 2010 30 40 3705 genes 123123 Normoxia Hypoxia 1 23123 Normoxia Hypoxia 3 123123 Normoxia Hypoxia z-score B 30 Principal component 3 (12.6%) 0-10 Generation-20 20 10 1stHypoxia 1stNormoxia 2ndHypoxia 2ndNormoxia 3rdHypoxia 3rdNormoxia 4thHypoxia 4thNormoxia C D Transgenerational hypoxia up-regulated 2222 2027 2 4 781 708 F Adjusted P value 0.01 0.02 0.03 Nervous system development Generation of neurons Neurogenesis Neuron differentiation Neuron development Neuron projection development Epidermis development 0.04 0 2 4 6 8 Count Plasma membrane bounded cell projection organization Cell projection organization Neuron projection morphogenesis Plasma membrane bounded cell projection morphogenesis Epidermal cell differentiation Cell projection morphogenesis Cell part morphogenesis Cell morphogenesis Epithelial cell differentiation Dendrite development Negative regulation of trans by RNA pol II Cellular component morphogenesis Cell fate commitment-30 Transgenerational hypoxia down-regulated Generation 1 2 3 4 1483 1063 598 328 Adjusted P value 5E-5 1E-4 0 10 20 30 Count Fig. 3. Repeated generational hypoxia exposure reveals transgenerationally adapted gene expression. (A and B) Repeated generational exposure to hypoxia in duces the dysregulation of a roughly similar set of genes at each generation as visualized by (A) heatmap and (B) principal components analysis. (C and D) venn diagrams reveal which transcripts are consistently (c) up- regulated or (d) down- regulated in response to repeated hypoxia exposure. (E) GO analysis of 73 consistently up- regulated genes in response to repeated hypoxia exposure in the first to third generation that are not up- regulated in the fourth generation reveals genes involved in stress re sponse, metabolism, apoptosis signaling, as well as lipid oxidation and transport. (F) GO analysis of 270 consistently down- regulated genes in response to repeated hy poxia exposure in the first to third generation that are not down- regulated in the fourth generation reveals genes involved in development, organization, and cell fate determination. treatment after one generation but not after two, three, or four gen erations as these would be hypoxia- responsive genes that adapted to regulate longevity; genes that were dysregulated in response to hy poxia treatment after one, two, and three generations but not after four generations would be hypoxia- responsive genes that adapted to regulate fertility. We found that, of the 3705 statistically significantly dysregulated genes in response to one generation of hypoxia exposure, 2027 genes were consistently up- regulated and 1063 genes were consistently down- regulated after two successive hypoxia exposures. Of these genes, 781 were consistently up- regulated and 598 were consistently down- regulated after three successive hypoxia exposures. Of these genes, 708 were consistently up- regulated and 328 were consistent ly down- regulated after four successive generations of hypoxia exposure (Fig. 3, C and D, and fig. S1C). We therefore focused on genes that were dysregulated when phenotypes of extended life span or reduced fertility were present but no longer dysregulated when those phenotypes disappeared (in the second and fourth generation, respectively). These comparisons revealed 195 up- regulated and 420 down- regulated genes that were dysregulated after one generation of hypoxia but not after two generations of successive hypoxia whose adapted expression are presumably involved in the intergenerational adaptation to hypoxia that regulates longevity. The adaptation genes were enriched for genes involved in sugar metabolism and immune response genes, which could potentially help to explain the extended longevity phenotype in the first but not the second generation (fig. S2, A and B). The comparisons of hypoxia- dysregulated genes in generations one, two, and three to those no longer dysregulated in 40 20 0-20-40 Principal component 2 (18.6%)
generation four (the transgenerational hypoxia- adapted fertility genes) consisted of 73 up- regulated genes (Fig. 3E) and 270 down- regulated genes of interest (Fig. 3F). An analysis of these transgen erationally adapted genes revealed up- regulated genes that were enriched in apoptosis signaling, the response to different metals, as well as lipid oxidation and transport (Fig. 3E). The down- regulated transgenerationally adapted genes were enriched in development, organization, and cell fate determination genes (Fig. 3F), which might help to explain the absence of a fertility defect in the fourth generation of successive hypoxia treatments. Together, these gene expression analyses revealed that most hypoxia- responsive genes still respond to hypoxia treatment despite repeated hypoxia expo sures (Fig. 3, A and B, and fig. S1B) and that only a very small num ber of genes displayed adaptations in terms of expression that could correlate with the adaptations of phenotypes that we observed. To further refine our examination of how transcription changes differ in a single generation exposure to hypoxia from repeated hy poxia exposures, we performed comparisons of the gene expression dataset generated in this study to the one we had previously gener ated (24). We found that the greater majority of the 949 shared hypoxia- responsive genes present in both datasets maintained con sistent dysregulation after repeated hypoxia exposure but mostly lost their dysregulation in subsequent generations in response to a single hypoxia exposure (fig. S3A). Analysis of this restricted set of hypoxia transgenerationally dysregulated genes did not reveal any major differences to the first dataset. A gene set enrichment analysis of the 134 genes that were dysregulated in the P0 but not in the F1 in response to repeated hypoxia did not reveal any obvious longevity regulating candidates (fig. S3B), although perhaps pathways in volved in protein and fat binding or lipid metabolism could be in volved in that hypoxia- adapted phenotype as lipid metabolism genes have been demonstrated to play a role in regulating longevity (30, 31). A gene ontology (GO) analysis of the 219 significantly dys regulated genes from the first, second, and third generations of re peated hypoxia that were not dysregulated in the fourth generation again revealed genes involved in development, which could explain the defects in fertility (fig. S3C). Together, these analyses begin to help to reveal how C. elegans can adapt to repeated generational stresses on a transcriptional level.
H3K27 trimethylase regulating enzymes are required for transgenerational adaptation
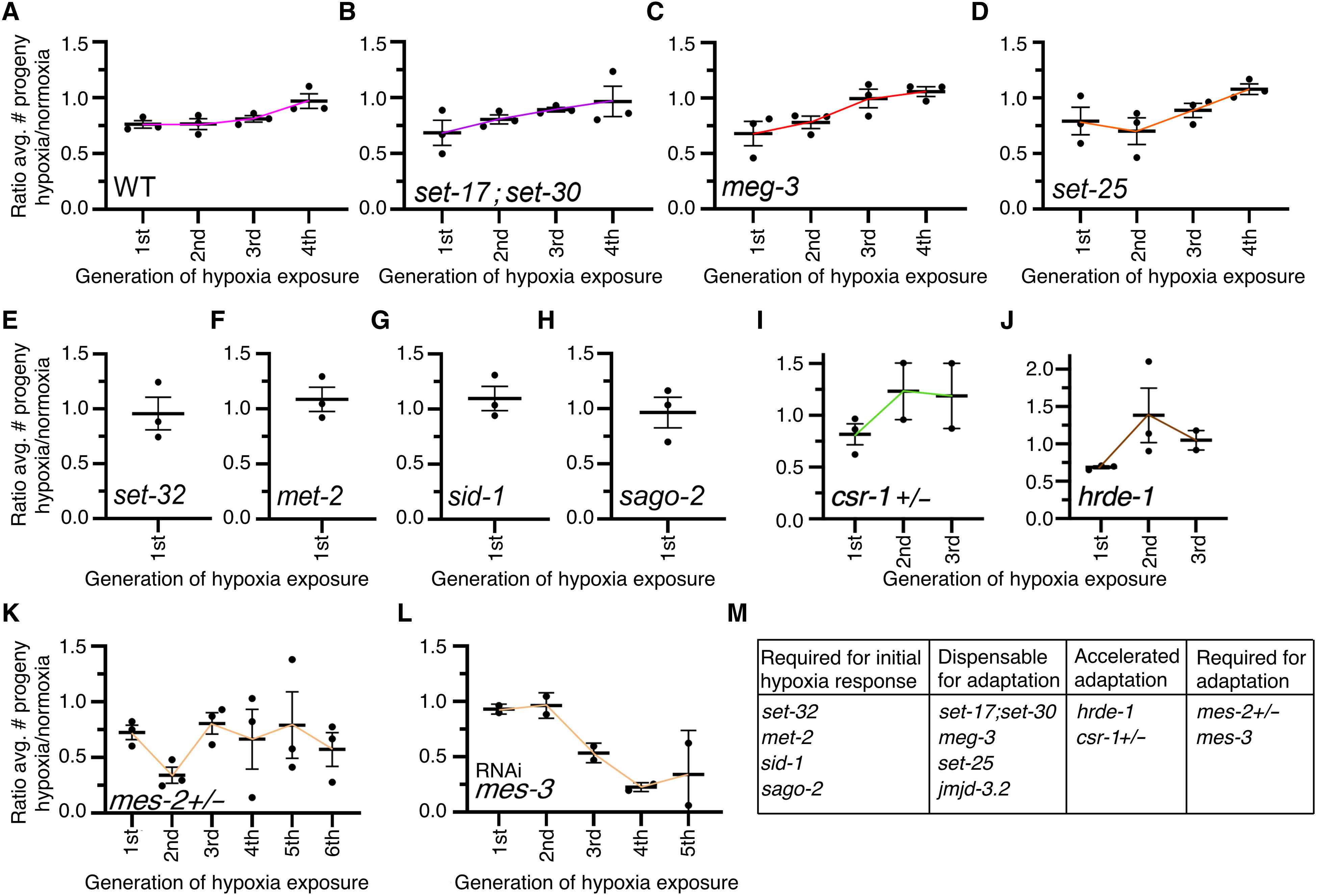
To determine how transgenerational adaptation to hypoxia was reg ulated, we first examined our RNA-seq data to see whether there were any epigenetic regulatory enzymes that displayed expression patterns that would match the phenotypic observations. However, we failed to observe any epigenetic regulators that displayed dys regulation after one generation of hypoxia but not subsequent gen erations or after three repeated generations of hypoxia but not after the fourth (fig. S2C). Because altered expression of epigenetic regu latory enzymes did not correlate with the transgenerational adapta tion phenotypes and because epigenetic regulatory enzymes are regulated in many different ways other than gene expression levels (32, 33), we proceeded to perform a directed genetic screen on epi genetic regulating genes that we had previously tested for their re quirements for transgenerational epigenetic inheritance in response to a single exposure to hypoxia (24). We identified that the H3K4 mono- and dimethyltransferases SET- 17 and SET- 30 (34), the in trinsically disordered protein MEG- 3, which is involved in phase separation through RNA binding (35), the putative H3K27me3 de methylase JMJD- 3.2 (36), and the putative H3K9 trimethyltransfer ase SET- 25 (37) were all dispensable for transgenerational adaptation to hypoxia (Fig. 4, A to D, and fig. S3). We had previously deter mined that jmjd- 3.2, set- 25, and meg- 3 were both required for the transgenerational epigenetic inheritance of fertility defects in re sponse to a single exposure of hypoxia (24). These genes are neces sary for the transgenerational hypoxia response but are dispensable for transgenerational adaptation, demonstrating that these disparate phenomena are regulated by independent mechanisms. As we had previously reported (24), we confirmed that the H3K23 trimethyltransferase SET- 32 (38), the dsRNA transporter SID- 1 (39), and the small RNA argonaute SAGO- 2 (40) were all re quired for the initial response to hypoxia and expanded these find ings by identifying that the putative H3K9 methyltransferase MET- 2 (41) was also required for the initial response to hypoxia (Fig. 4, E to H), and therefore it is not possible to determine whether these genes play a role in transgenerational adaptation. We determined that de letion of the argonaute proteins, CSR- 1 (42) and HRDE- 1 (43), ac celerated the adaptation to hypoxia (Fig. 4, I and J). Although the hypoxia- exposed csr- 1 and hrde- 1 mutant worms laid fewer eggs in the first generation, after another successive exposure to hypoxia, csr- 1 and hrde- 1 mutant worms laid comparable or even slightly higher numbers of eggs to normoxia worms (Fig. 4, I and J). We had previously found that hrde- 1 was required for transmitting the transgenerational epigenetic inheritance of fertility defects in re sponse to a single exposure of hypoxia (24), suggesting that small RNAs are involved in both the transgenerational hypoxia response and the transgenerational adaptation process. We identified that worms that were heterozygous for the puta tive H3K27 trimethyltransferase mes- 2 (44) never adapted to repeated successive hypoxia exposures (Fig. 4K). MES- 2 is the catalytic enzyme of a multisubunit protein complex, termed the polycomb repressive complex 2 (PRC2), that is important for maintaining repressive H3K 27me3 chromatin modifications (45–48). We had previously found that mes- 2 was also required for inheritance in response to a single generation exposure to hypoxia (24). MES- 3 is an ortholog of the sup pressor of Zeste 12 (SUZ12) (44, 49), a core subunit of the PRC2 complex that provides a structural scaffold for the complex and is necessary for the methyltransferase activity (50, 51). mes- 3 knockdown also pre vented adaptation to repeated successive hypoxia exposures (Fig. 4L). mes- 3 knockdown in conjunction with hypoxia treatment made the fertility relative to normoxia virtually indistinguishable until the third generation. This finding could be due to incomplete penetrance of RNA interference (RNAi), a resetting of exogenous heritable dsRNA (52), or a synergism between the maternal effect sterile phenotype of mes- 3 depletion and hypoxia treatment having altered penetrance over successive generations. Together, however, these results suggest that this complex and H3K27 methylation are required for the trans generational adaptation to hypoxia.
DISCUSSION
Through these experiments, we showed that C. elegans adapt on a transgenerational scale to repeated hypoxia and elevated glucose ex posures. We found that, after two generations of successive hypoxia exposure, C. elegans no longer display a hypoxia- induced life- span ex tension (Fig. 1D). We also found that, after four generations of succes sive hypoxia exposure, C. elegans no longer display a hypoxia- induced
Fig. 4. The PRC2 complex is required for transgenerational adaptation to hypoxia. (A to D) set- 17;set- 30, meg- 3, and set- 25 mutant worms all adapted to repeated generational hypoxia exposure after four generations as Wt worms do, suggesting that these genes are not involved in transgenerational adaptation to repeated hy poxia exposure. (E to H) set- 32, met- 2, sid- 1, and sago- 2 mutant worms do not display reduced fertility in response to a single generation of hypoxia, suggesting that these genes are necessary for the initial hypoxia response. (I and J) csr- 1 heterozygous and hrde- 1 mutant worms adapt to repeated generational hypoxia exposure after two generations rather than four generations, suggesting that these argonaute genes are required for maintaining the generational clock. (K and L) mes- 2 heterozygous mutant worms and knockdown of mes- 3 eliminated the adaptation to repeated generational hypoxia exposure, suggesting that these genes are required for transgen erational adaptation. each dot represents an independent experiment consisted of three replicate plates with 10 worms per plate. For hrde- 1 and mes- 2 mutant strains, 20 worms were used per plate. the ratio of the average number of eggs laid by hypoxia- treated worms divided by the average number of eggs laid by normoxia- treated worms is displayed. error bars represent the SeM, and colored lines are used as a trendline across generations. (M) Summary of all genetic screening experiments. fertility defect (Fig. 1B). This phenomenon was not specific to hypoxia exposure as repeated generation exposure to elevated glucose also caused an adaptation in C. elegans (Fig. 2). We observed that, for the most part, repeated hypoxia exposure continued to cause the dysregu lation of many transcripts (Fig. 3A and fig. S1B). However, we identi fied a subset of transgenerationally adapted genes whose expression was no longer altered in response to two successive exposures to hy poxia and were involved in sugar metabolism and immune response genes, raising the possibility that these genes could be important for life- span regulation in response to hypoxia. Similarly, we identified a subset of transgenerationally adapted genes whose expression was no longer altered in response to four successive exposures to hypoxia and were involved in apoptosis signaling, the response to different metals, as well as lipid oxidation and transport, raising the possibility that these genes are important for fertility regulation. Through a directed genetic screen, we found that the argonautes CSR- 1 and HRDE- 1 as well as the H3K27 trimethyltransferase complex are necessary for transgenerational adaptation to hypoxia. Together, these results dem onstrate that organisms can adapt on a transgenerational scale to repeated stresses and that this transgenerational adaptation shares some of the same epigenetic regulating enzymes that are required for the initial transgenerational response to that same stress. Our previous work had identified that both HRDE- 1 and MES- 2 were necessary for the transgenerational response to a single gen erational exposure to hypoxia (24). When these genes were deleted, C. elegans displayed an initial response to hypoxia but were unable to transmit the phenotypes to naïve normoxia- reared descendants (24). Here, we found that hrde- 1 deletion and ablation of the PRC2 complex had opposite phenotypic consequences. Deletion of hrde- 1 accelerat ed the transgenerational adaptation, such that two successive genera tions of hypoxia exposure was sufficient to induce transgenerational adaptation instead of four repeated generations (Fig. 4J), whereas ab lation of the PRC2 complex made it so that worms were resistant to transgenerational adaptation (Fig. 4, K and L). Although these find ings are mostly phenomenological in their essence, they do point to some exciting potential mechanisms that could be further followed up on with regard to small RNAs and H3K27me3 inheritance. It is pos sible that one set of small RNAs could be responsible for an initial
transgenerational response whereas a second set of small RNAs could be responsible for the transgenerational adaptation response. Simi larly, H3K27me3 could mark the histones of one set of genes impor tant for an initial transgenerational response and a second set of genes important for the transgenerational adaptation response, and both sets of genes and small RNAs could independently be important for conferring extended longevity and reduced fertility. Many, but not all, of the genes we identified as being necessary for transgenerational epigenetic inheritance in response to a single ex posure of hypoxia (24) were also involved in the transgenerational adaptation to repeated hypoxia exposure. Because the transgenera tional epigenetic inheritance and the transgenerational adaptation to hypoxia are almost two opposing phenomena, it is perhaps expected that small RNA regulation is necessary for the former and deletion accelerates the latter. However, ablation of PRC2 inhibits both trans generational epigenetic inheritance and transgenerational adapta tion from occurring. This finding harkens back to the opposing role of EZH2 as an oncogene or a tumor suppressor (53–56) and a pro- aging and pro- longevity signal depending on its context (57–59), highlighting the context- dependent nature of epigenetic regulators. This reinforces that these enzymes may have alternative functions in different cell types or circumstances. It will be interesting, in future studies, to investigate how H3K27me3 regulation can accomplish these opposing actions in transgenerational epigenetic inheritance. Unexpectedly, the transgenerational adaptation of different traits, such as life span and fertility, took place on a different generational time scale, with longevity adapting after two successive generations but fertility requiring four to fully adapt. This difference suggests that either the two different traits have different thresholds for al tered phenotypes and there is simply a slow decrease in the heritable molecules, which happens to pass the threshold for longevity be fore the threshold for altering fertility occurs, or the environmental changes are inducing many independent signaling pathways in par allel and some are no longer induced in response to repeated hy poxia more rapidly than other signaling pathways. These two options are not mutually exclusive, and it will be interesting, in future experiments, to identify the universe of all heritable materials and identify which molecules are specifically responsible for which heritable phenotype. We had also previously identified that a single generation of hy poxia exposure caused an intergenerational reduction of neutral lip ids (24). In this current study, we found that our GO analysis for transgenerationally adapted up- regulated genes revealed an enrich ment for genes involved in lipid oxidation and lipid transport. Given that some lipids function as signaling molecules between cells (60) and between organisms (61) and even potentially across generations (62, 63), it will be interesting to determine whether altered lipid lev els are simply a consequence of hypoxia treatment or could be in volved in transmitting nongenetic information across generations. Transgenerational epigenetic inheritance has always functioned at the interface between genetics and environment, allowing organ isms to adapt on a relatively short time scale, from an evolutionary perspective, of a few generations. This phenomenon allows organ isms to inform the subsequent generations that conditions might not be optimal and they should therefore adapt to prepare for such an eventuality. Transgenerational adaptation, on the other hand, would exist if such a detrimental environmental condition were per petually present; the organism would not want to persist in a height ened adapted condition but would instead accept that these were the new conditions for the foreseeable future in their domain. It will be exciting to determine whether transgenerational adaptation is a prevalent phenomenon, occurring in response to many environ mental stressors that induce transgenerational epigenetic inheri tance, or whether this is a unique responder to hypoxia and glucose. If this phenomenon is present in response to multiple stresses, it will be interesting to investigate whether each instance of transgenera tional adaptation is dependent on small RNAs and H3K27me3 reg ulating enzymes or, if like transgenerational epigenetic inheritance itself, these transgenerational adaptations function through multi ple independent parallel epigenetic regulatory mechanisms.
MATERIALS AND METHODS
Experimental design
The objectives of this study are to understand how C. elegans re spond to repeated generational exposure to stressors such as hypoxia and glucose.
Hypoxia exposure
The worms were kept at 20°C and were well fed before exposure. L4 worms were transferred to a new Nematode Growth Media (NGM) plate with sufficient food and were exposed to a 0.1% oxygen lev el for 16 hours using an Eppendorf Galaxy 48R incubator. After 16 hours, they were moved back to normal atmospheric oxygen level (~20% O2). Normoxia/control worms remained in an incubator at normal oxygen levels for the entire experiment. The progeny of the first generation were selected at the L4 stage, and this process was repeated across multiple generations (Fig. 1A).
Glucose exposure
Glucose (1 or 2%) was added in NGM plates seeded with OP50. Eggs were put onto plates after bleaching and allowed to grow til day 1 of adulthood at 20°C. For subsequent generations, 20 to 30 worms were transferred onto fresh OP50 with or without glucose supplementation and allowed to lay eggs for 2 hours. The mother worms were then euthanized. In each generation, oil red O was performed at the post–young adult stage.
C. elegans strains
The N2 Bristol strain was used as the wild- type (WT) background. The following mutations were used in this study: hrde- 1(tm1200), csr- 1(tm892) IV/nT1 [unc- ?(n754) let- ?] (IV;V), mes- 3(bn21), jmjd- 3.1(gk384), jmjd- 3.1(gk387), jmjd- 3.2(tm3121), meg- 3(tm4259), mes- 2(bn11) unc- 4(e120)/mnC1 [dpy- 10(e128) unc- 52(e444)], met- 2(ok2307), sago- 2(tm894), set- 17(n5017)/set- 30(gk315), set- 25 (n5021), set- 32(ok1457), and sid- 1(qt9). Worms were grown on OP50 bacteria in all experiments.
Antibodies
Antibodies used for CUT&RUN were rabbit anti- H3 (Abcam, Ab1791) and rabbit anti- H3K27me3 (MilliporeSigma, 07- 449). These antibodies have demonstrated specificity in eukaryotes. The CUT&RUN ex periment was unsuccessful due to technical difficulties.
Fertility assay
NGM plates with OP50 were prepared as described above. Ten worms at the L4 stage were transferred to OP50- seeded NGM plates in trip licate (30 worms per condition) and kept at 20°C. The worms were transferred to new plates every day for 2 days. The number of hatched worms was counted.
Mes- 3 RNAi fertility assay
Preparation of mes- 3 RNAi petri plates: 20 g of agar, 3 g of NaCl, and 5 g of Bacto Peptone was added to a 2- liter flask with 1 liter of deionized H2O and autoclaved for 55 min on liquid cycle. The flask was cooled to ~55°C before adding in 25 ml of 1 M KPO4 buffer, 1 ml of 1 M MgSO4 buffer, 1 ml of 1 M CaCl2 buffer, 1 ml of cholesterol (5 mg/ml) dissolved in ethanol, 0.4 mM IPTG (isopropyl- β- d- thiogalactopyranoside), and 0.5 ml of ampicillin (3 mg/ml). Ten milliliters of this mixture was add ed to each petri dish (60 mm by 15 mm) and left at room temperature for 24 hours to solidify. Two hundred microliters of F54C1.3 bacteria was added for the mes- 3 plates and HT115(DE3) bacteria for the empty vector control. L4 worms were transferred from OP50 NGM plates to their respective plates. The hypoxia exposure step and fertility assay were conducted as described above.
Longevity assay
Worm life- span assays were conducted at 20°C, and worms were grown on OP50 NGM plates without 5- fluoro- 2′- deoxyuridine (FUdR). For each assay, roughly 90 worms per condition were ob served on three plates (30 worms per plate) at the beginning of the experiment. Worms that crawled off the plate, had a ruptured vulva, or experienced matricide were censored. Kaplan- Meier curves were plotted and visualized in GraphPad Prism. P values were calculated using log- rank tests.
RNA sequencing
C. elegans age synchronization
Plates with worms were synchronized by washing the adult worms off the plates using M9 buffer with 1% Triton X- 100 once a sufficient number of eggs were laid. Through gentle pipetting, the worms are washed away whereas the eggs remain intact on the bacterial lawn. These eggs were reared at 20°C and hatched. Once most worms ap peared to be at the L4 stage, they were exposed to either normoxia or hypoxia as per the experimental setup.
C. elegans sample collection
Because this experiment focuses on multiple generations of C. elegans, it was important to collect and freeze worm samples so that they could be used for the following experiments simultane ously. Three biological replicates for each condition between the first and fourth generations were produced, and a total of 24 samples were collected. After the 16- hour exposure to normoxia or hypoxia, worms were washed off the plate using M9 buffer with 1% Triton X- 100 and collected in a 1.5- ml microcentrifuge tube. The worms were allowed to settle to the bottom, and the excess buffer was re moved. The worm pellet was then flash frozen in liquid nitrogen for 5 s and stored at −80°C until use.
Total RNA extraction for RNA-seq
Total RNA was extracted using the Direct-zol RNA Miniprep Kit from Zymo Research. Briefly, frozen worm samples were lysed by adding in 300 μl of TRI Reagent and homogenizing with a Kontes pellet pestle motor for 1 min. An equal amount of ethanol was add ed and mixed thoroughly before being transferred to a Zymo- Spin IICR Column in a collection tube and centrifuged. The column ma trix was treated with DNase I and DNA Digestion Buffer and incu bated at room temperature for 15 min to get rid of any unwanted traces of DNA. Afterward, the column was washed twice with Direct-zol RNA PreWash and once with RNA Wash Buffer. The col umn was then transferred in a clean tube, and the RNA was eluted by adding 25 μl of DNase/RNase- Free Water to the column matrix and centrifuged for 30 s. The RNA concentration and purity were measured using a NanoDrop before storing at −80°C until use.
RNA-seq library preparation
- mRNA Enrichment
- mRNA was enriched from total RNA samples using NEXTFLEX Poly(A) Beads 2.0.
- Library Preparation
- Libraries for RNA-seq with these samples were prepared using the NEXTFLEX Rapid Directional RNA-Seq Kit 2.0.
- Twenty-four samples were prepared, each with 1 μg of the mRNA starting material.
- In short, mRNA is fragmented before first and second strand synthesis.
- Strands are then adenylated, and index adapters are ligated, followed by 11 cycles of polymerase chain reaction (PCR) amplification.
- The PCR products were cleaned up using NEXTFLEX Cleanup Beads XP and transferred to nonstick RNase-free microcentrifuge tubes and stored at −20°C until sample submission.
- RNA Validation
- RNA quality was validated using Agilent 2200 TapeStation D1000 before being sequenced using a NovaSeq 6000 SP platform. Fastq files were checked for quality using FastQC before being filtered out with a Q score of 30 or above and trimmed for adapters using CutAdapt 4.0. The trimmed reads were then aligned to the C. elegans genome (WS235) using bowtie2, and samtools was used to convert sam outputs to sorted bam files. FeatureCounts was used to quantify the number of counts for each gene using the sorted bam files. Genes under 350 counts were filtered out, and DESeq2 was used to normalize raw counts. Differentially expressed genes with a P value of > 0.05 was filtered out before downstream analyses. RNA-seq figures were plotted using ggplot2, ComplexHeatMap, clusterProfiler, AnnotationDbi, org.Ce.eg.db, and EnhancedVolcano packages in RStudio.
Oil red O staining
- Egg Growth
- Eggs were grown in NGM OP50 with or without 1 or 2% glucose addition.
- Staining
- Day 1 adults were stained with oil red O as previously described (64).
- Mounting
- Worms were mounted on a glass slide with an agarose pad [2% (w/v) agarose in ddH2O] and visualized under a bright-field microscope (Leica Dmi8).
- The entire worm oil red O staining was quantified using ImageJ Fiji 5.
Statistical analysis
The number of replicates, P values, and statistical test used for each experiment is indicated in the corresponding figure legend and detailed in this section. P values for independent replicates of fertility assays (Fig. 1B) were determined by two-tailed t test. P values for Kaplan- Meier curves (Fig. 1, C and D) were calculated using log- rank rests (Mantel- Cox), and P values for glucose exposure assays (Fig. 2, A, C, E, G, H, and I) were determined using one- way analy sis of variance (ANOVA). P values for independent replicates were combined using combined Fisher’s test. The r2 value quantifying the relationship between the logFC (log fold change) values of signifi cant genes in the P0 generation for this study and previously ob served differentially expressed genes from hypoxia treatment (24) was derived from a standard linear regression model (fig. S1A).
Figures
Figure 1.
...
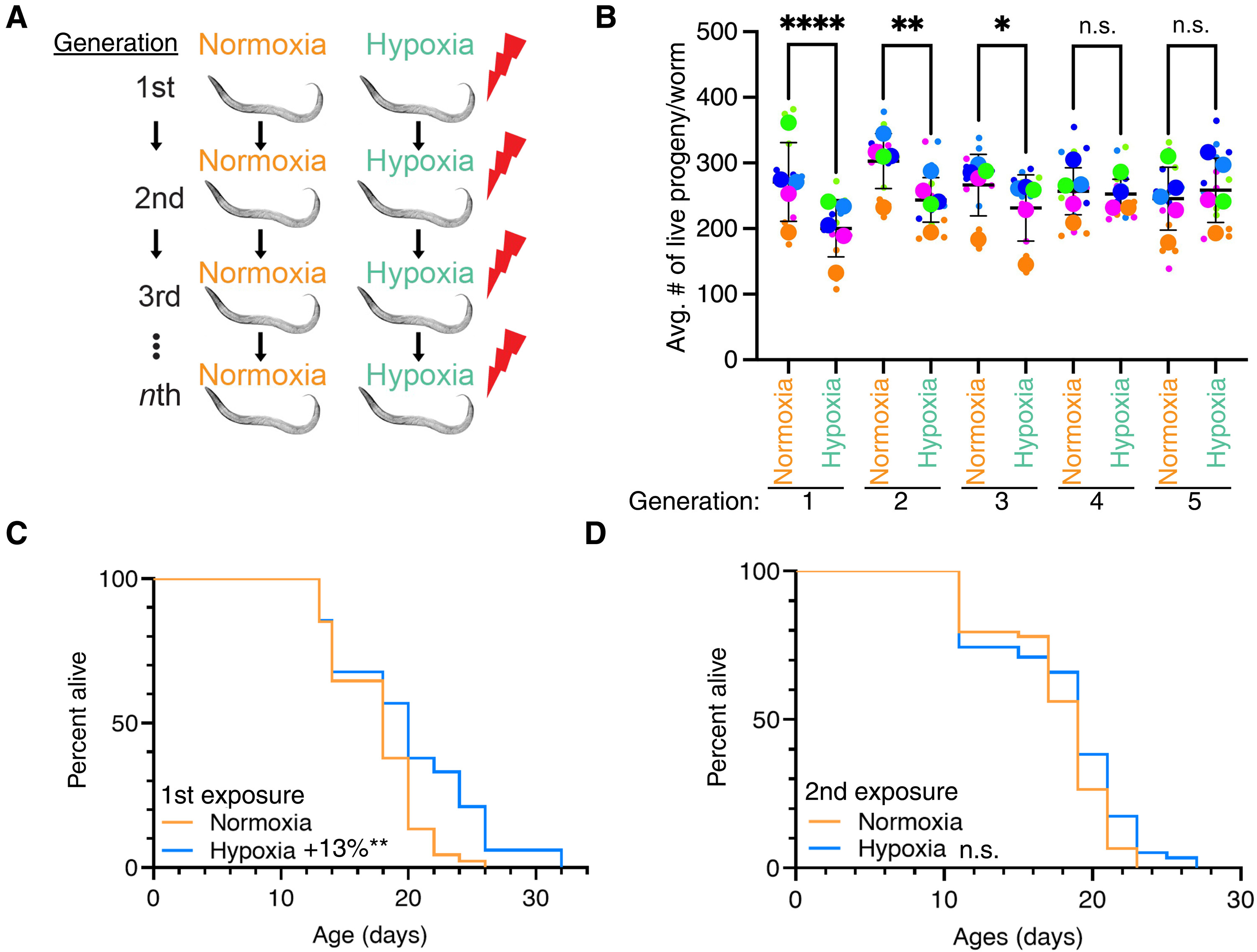
Figure 2.
...
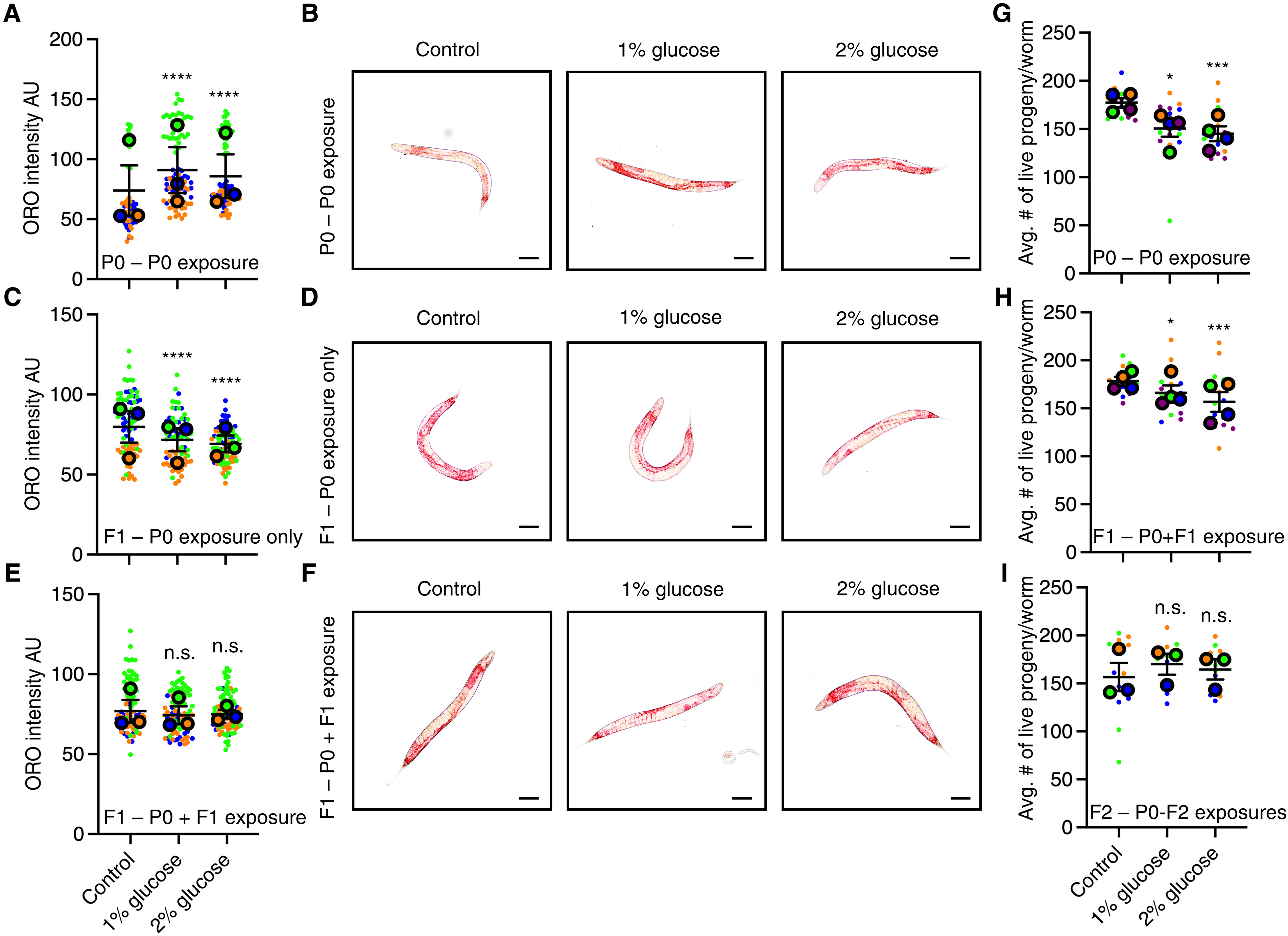
Figure 3.
...
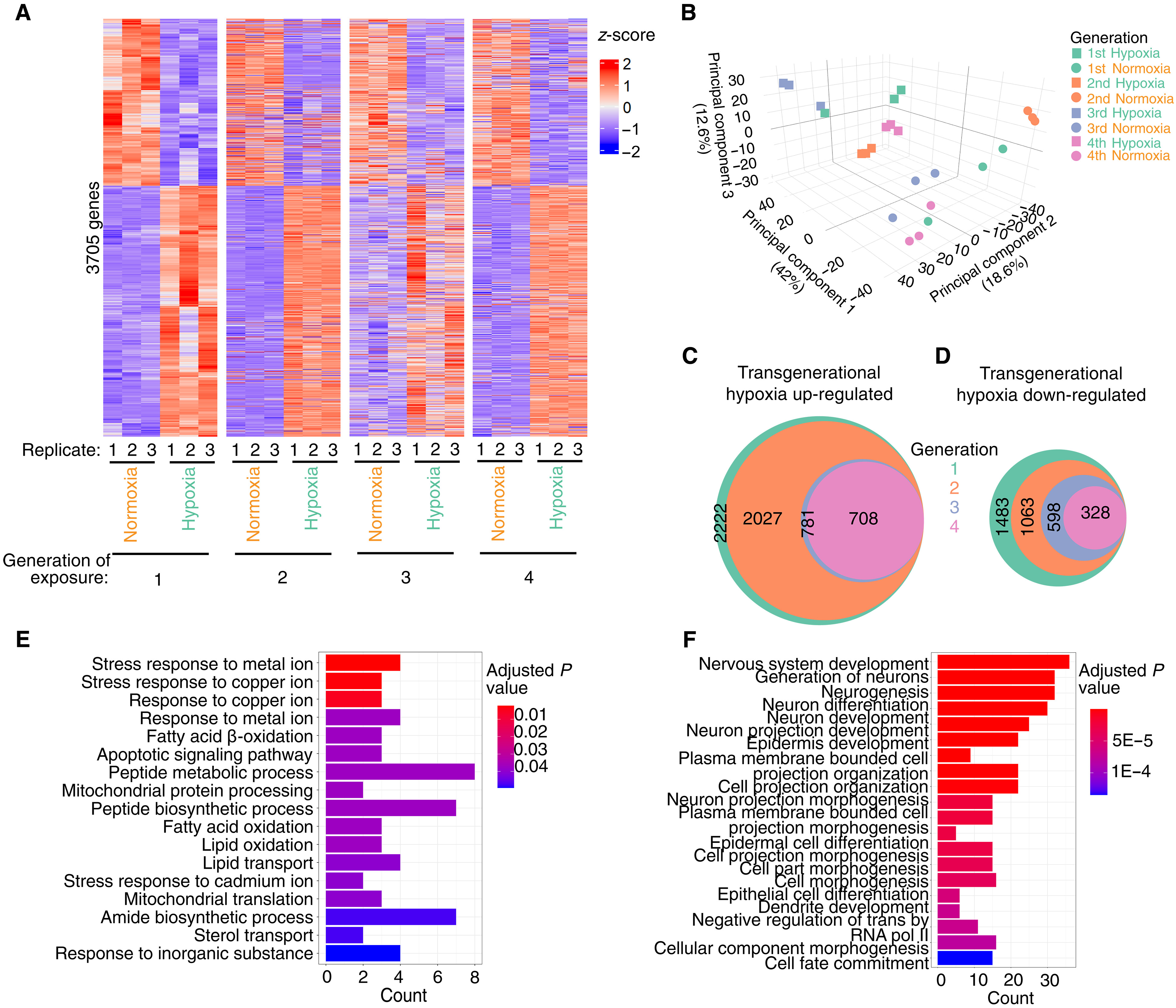
Figure 4.
...
Avg.#of live progeny/worm C D 1st exposure Normoxia Hypoxia Normoxia Hypoxia Normoxia Hypoxia Normoxia Hypoxia Normoxia Hypoxia Generation: +13%** 1 3 2 4 5 2nd exposure n.s. Fig. 1. C. elegans adapt transgenerationally to repeated hypoxia exposure. (A) Scheme of the repeated generational hypoxia exposure. (B) Repeated generational hypoxia exposure leads to decreased fertility for three successive generations but fails to induce a significant fertility defect after four repeated exposures. each color- coded dot represents an individual experiment performed in triplicate with 10 worms per plate, and each column represents the means ± SeM. *P < 0.05; **P < 0.01; ****P < 0.0001; not significant (n.s.), P > 0.05, as assessed by Fisher’s combined probability test. data are presented in superplot format (65) with the average of each ex periment shown in a larger colored dot. (C and D) hypoxia exposure induces life- span extension after one exposure (c) but not after two generational exposures (d). Statistics are presented in table S1.
A C
P0 – P0 exposure
B P0 – P0 exposure D F1 – P0 exposure only E F n.s. n.s. F1 – P0 + F1 exposure F1 – P0 exposure only F1 – P0 + F1 exposure G Control 1% glucose 2 % glucose Control 1% glucose 2% glucose H I Control 1% glucose 2% glucose * Avg. # of live progeny/worm
P0 – P0 exposure * Avg. # of live progeny/worm
F1 – P0+F1 exposure n.s. n.s. Avg. # of live progeny/worm F2 –P0-F2 exposures Fig. 2. C. elegans adapt transgenerationally to repeated elevated glucose exposure. (A and B) exposure to 1 or 2% glucose leads to elevated neutral lipid levels as assessed by oil red O (ORO) staining in the parental P0 generation. (A) each large dot represents replicate experiments performed with 13 to 34 worms. AU, arbitrary units. (B) Represen tative images of the P0 exposed generation. (C and D) exposure to 1 or 2% glucose in parental generation leads to decreased levels of neutral lipids in the F1 unexposed progeny by ORO staining. (c) each large dot represents replicate experiments performed with 13 to 38 worms. (d) Representative images of the F1 progeny. (E and F) Repeated exposure to 1 or 2% glucose in the parental and F1 generation eliminates the increase in neutral lipids observed in the P0 generation as assessed by ORO staining. (e) each large dot represents replicate experiments performed with 18 to 55 worms. (F) Representative images of the F1 progeny exposed to different amounts of glucose, whose parents were also exposed to those altered glucose levels. (G) exposure to 1 or 2% glucose leads to decreased fertility. each large dot represents an individual experiment performed in triplicate with 10 worms per plate. (H) two successive generational exposures to 1 or 2% glucose lead to decreased fertility. (I) three successive generational exposures to 1 or 2% glucose fail to induce decreased fertility. in all graphs, each color- coded dot represents the means ± SeM with individual replicates within an experiment presented in the same color with a smaller dot in superplot format (63). *P < 0.05; ***P < 0.001; ****P < 0.0001; ns, P > 0.05, as assessed by Fisher’s combined probability test.
REFERENCES AND NOTES
- O. t. Avery, c. M. Macleod, M. Mccarty, Studies on the chemical nature of the substance inducing transformation of pneumococcal types: induction of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type iii. J. Exp. Med. 79, 137–158 (1944).
- h. d. Morgan, h. G. Sutherland, d. i. Martin, e. Whitelaw, epigenetic inheritance at the agouti locus in the mouse. Nat. Genet. 23, 314–318 (1999).
- G. cavalli, R. Paro, the drosophila Fab- 7 chromosomal element conveys epigenetic inheritance during mitosis and meiosis. Cell 93, 505–518 (1998).
- d. c. Benyshek, c. S. Johnston, J. F. Martin, Glucose metabolism is altered in the adequately- nourished grand- offspring (F3 generation) of rats malnourished during gestation and perinatal life. Diabetologia 49, 1117–1119 (2006).
- B. G. dias, K. J. Ressler, Parental olfactory experience influences behavior and neural structure in subsequent generations. Nat. Neurosci. 17, 89–96 (2014).
- e. l. Greer, t. J. Maures, d. Ucar, A. G. hauswirth, e. Mancini, J. P. lim, B. A. Benayoun, Y. Shi, A. Brunet, transgenerational epigenetic inheritance of longevity in Caenorhabditis elegans. Nature 479, 365–371 (2011).
- e. l. Greer, B. Becker, c. latza, A. Antebi, Y. Shi, Mutation of C. elegans demethylase spr- 5 extends transgenerational longevity. Cell Res. 26, 229–238 (2016).
- M. h. Rothi, e. l. Greer, From correlation to causation: the new frontier of transgenerational epigenetic inheritance. Bioessays 45, e2200118 (2023).
- c. h. Waddington, canalization of development and the inheritance of acquired characters. Nature 150, 563–565 (1942).
- G. loor, P. t. Schumacker, Role of hypoxia- inducible factor in cell survival during myocardial ischemia- reperfusion. Cell Death Differ. 15, 686–690 (2008).
- G. l. Semenza, the hypoxic tumor microenvironment: A driving force for breast cancer progression. Biochim. Biophys. Acta 1863, 382–391 (2016).
- l. Fajersztajn, M. M. veras, hypoxia: From placental development to fetal programming. Birth Defects Res. 109, 1377–1385 (2017).
- S. Gurusinghe, A. tambay, c. B. Sethna, developmental origins and nephron endowment in hypertension. Front. Pediatr. 5, 151 (2017).
- c. Kanaka- Gantenbein, Fetal origins of adult diabetes. Ann. N. Y. Acad. Sci. 1205, 99–105 (2010).
- i. c. McMillen, J. S. Robinson, developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 85, 571–633 (2005).
- R. J. diaz, R. Rosenberg, Spreading dead zones and consequences for marine ecosystems. Science 321, 926–929 (2008).
- W. ni, M. li, A. c. Ross, R. G. najjar, large projected decline in dissolved oxygen in a eutrophic estuary due to climate change. J. Geophys. Res. Oceans 124, 8271–8289 (2019).
- J. Zhang, G. cowie, S. W. A. naqvi, hypoxia in the changing marine environment.
Environ. Res. Lett. 8, 015025 (2013). - A. A. chakraborty, t. laukka, M. Myllykoski, A. e. Ringel, M. A. Booker, M. Y. tolstorukov,
Y. J. Meng, S. R. Meier, R. B. Jennings, A. l. creech, Z. t. herbert, S. K. McBrayer,
B. A. Olenchock, J. d. Jaffe, M. c. haigis, R. Beroukhim, S. Signoretti, P. Koivunen,
W. G. Kaelin, histone demethylase KdM6A directly senses oxygen to control chromatin and cell fate. Science 363, 1217–1222 (2019). - W. A. van voorhies, S. Ward, Broad oxygen tolerance in the nematode Caenorhabditis elegans. J. Exp. Biol. 203, 2467–2478 (2000).
- t. heimbucher, J. hog, P. Gupta, c. t. Murphy, PQM- 1 controls hypoxic survival via regulation of lipid metabolism. Nat. Commun. 11, 4627 (2020).
- S. F. leiser, M. Fletcher, A. Begun, M. Kaeberlein, life- span extension from hypoxia in
Caenorhabditis elegans requires both hiF- 1 and dAF- 16 and is antagonized by SKn- 1.
J. Gerontol. A Biol. Sci. Med. Sci. 68, 1135–1144 (2013). - c. l. Pender, h. R. horvitz, hypoxia- inducible factor cell non- autonomously regulates
C. elegans stress responses and behavior via a nuclear receptor. eLife 7, e36828 (2018). - S. Y. Wang, K. Kim, Z. K. O’Brown, A. levan, A. e. dodson, S. G. Kennedy, c. chernoff,
e. l. Greer, hypoxia induces transgenerational epigenetic inheritance of small RnAs.
Cell Rep. 41, 111800 (2022). - h. Adachi, Y. Fujiwara, n. ishii, effects of oxygen on protein carbonyl and aging in
Caenorhabditis elegans mutants with long (age- 1) and short (mev- 1) life spans.
J. Gerontol. A Biol. Sci. Med. Sci. 53, B240–B244 (1998). - A. tauffenberger, A. vaccaro, A. Aulas, c. vande velde, J. A. Parker, Glucose delays age- dependent proteotoxicity. Aging Cell 11, 856–866 (2012).
- S. J. lee, c. t. Murphy, c. Kenyon, Glucose shortens the life span of C. elegans by downregulating dAF- 16/FOXO activity and aquaporin gene expression. Cell Metab. 10, 379–391 (2009).
- M. A. Mondoux, d. c. love, S. K. Ghosh, t. Fukushige, M. Bond, G. R. Weerasinghe,
J. A. hanover, M. W. Krause, O- linked- n- acetylglucosamine cycling and insulin signaling are required for the glucose stress response in Caenorhabditis elegans. Genetics 188, 369–382 (2011). - A. tauffenberger, J. A. Parker, heritable transmission of stress resistance by high dietary glucose in Caenorhabditis elegans. PLOS Genet. 10, e1004346 (2014).
- M. c. Wang, e. J. O’Rourke, G. Ruvkun, Fat metabolism links germline stem cells and longevity in C. elegans. Science 322, 957–960 (2008).
- l. R. lapierre, S. Gelino, A. Meléndez, M. hansen, Autophagy and lipid metabolism coordinately modulate life span in germline- less C. elegans. Curr. Biol. 21, 1507–1514 (2011).
- K. tzelepis, O. Rausch, t. Kouzarides, RnA- modifying enzymes and their function in a chromatin context. Nat. Struct. Mol. Biol. 26, 858–862 (2019).
- Y. nie, c. Song, h. huang, S. Mao, K. ding, h. tang, chromatin modifiers in human disease: From functional roles to regulatory mechanisms. Mol. Biomed. 5, 12 (2024).
- e. l. Greer, S. e. Beese- Sims, e. Brookes, R. Spadafora, Y. Zhu, S. B. Rothbart,
d. Aristizábal- corrales, S. chen, A. i. Badeaux, Q. Jin, W. Wang, B. d. Strahl, M. P. colaiácovo, Y. Shi, A histone methylation network regulates transgenerational epigenetic memory in c. elegans. Cell Rep. 7, 113–126 (2014). - J. Smith, d. calidas, h. Schmidt, t. lu, d. Rasoloson, G. Seydoux, Spatial patterning of P granules by RnA- induced phase separation of the intrinsically- disordered protein MeG- 3. eLife 5, e21337 (2016).
- J. vandamme, G. lettier, S. Sidoli, e. di Schiavi, O. nørregaard Jensen, A. e. Salcini, the C. elegans h3K27 demethylase UtX- 1 is essential for normal development, independent of its enzymatic activity. PLOS Genet. 8, e1002647 (2012).
- B. d. towbin, c. González- Aguilera, R. Sack, d. Gaidatzis, v. Kalck, P. Meister, P. Askjaer,
S. M. Gasser, Step- wise methylation of histone h3K9 positions heterochromatin at the nuclear periphery. Cell 150, 934–947 (2012). - l. Schwartz- Orbach, c. Zhang, S. Sidoli, R. Amin, d. Kaur, A. Zhebrun, J. ni, S. G. Gu, Caenorhabditis elegans nuclear RnAi factor Set- 32 deposits the transgenerational histone modification, h3K23me3. eLife 9, e54309 (2020).
- A. M. Jose, J. J. Smith, c. P. hunter, export of RnA silencing from C. elegans tissues does not require the RnA channel Sid- 1. Proc. Natl. Acad. Sci. U.S.A. 106, 2283–2288 (2009).
- e. Yigit, P. J. Batista, Y. Bei, K. M. Pang, c. c. chen, n. h. tolia, l. Joshua- tor, S. Mitani,
M. J. Simard, c. c. Mello, Analysis of the c. elegans Argonaute family reveals that distinct Argonautes act sequentially during RnAi. Cell 127, 747–757 (2006). - e. c. Andersen, h. R. horvitz, two c. elegans histone methyltransferases repress lin- 3 eGF transcription to inhibit vulval development. Development 134, 2991–2999 (2007).
- J. M. claycomb, P. J. Batista, K. M. Pang, W. Gu, J. J. vasale, J. c. van Wolfswinkel,
d. A. chaves, M. Shirayama, S. Mitani, R. F. Ketting, d. conte Jr., c. c. Mello, the Argonaute cSR- 1 and its 22G- RnA cofactors are required for holocentric chromosome segregation. Cell 139, 123–134 (2009). - B. A. Buckley, K. B. Burkhart, S. G. Gu, G. Spracklin, A. Kershner, h. Fritz, J. Kimble, A. Fire,
S. Kennedy, A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature 489, 447–451 (2012). - l. B. Bender, R. cao, Y. Zhang, S. Strome, the MeS- 2/MeS- 3/MeS- 6 complex and regulation of histone h3 methylation in C. elegans. Curr. Biol. 14, 1639–1643 (2004).
- R. cao, l. Wang, h. Wang, l. Xia, h. erdjument- Bromage, P. tempst, R. S. Jones, Y. Zhang, Role of histone h3 lysine 27 methylation in Polycomb- group silencing. Science 298, 1039–1043 (2002).
- B. czermin, R. Melfi, d. Mccabe, v. Seitz, A. imhof, v. Pirrotta, drosophila enhancer of Zeste/eSc complexes have a histone h3 methyltransferase activity that marks chromosomal Polycomb sites. Cell 111, 185–196 (2002).
- A. Kuzmichev, K. nishioka, h. erdjument- Bromage, P. tempst, d. Reinberg, histone methyltransferase activity associated with a human multiprotein complex containing the enhancer of Zeste protein. Genes Dev. 16, 2893–2905 (2002).
- J. Müller, c. M. hart, n. J. Francis, M. l. vargas, A. Sengupta, B. Wild, e. l. Miller,
M. B. O’connor, R. e. Kingston, J. A. Simon, histone methyltransferase activity of a drosophila Polycomb group repressor complex. Cell 111, 197–208 (2002). - B. Snel, S. van den heuvel, M. F. Seidl, Caenorhabditis elegans MeS- 3 is a highly divergent ortholog of the canonical PRc2 component SUZ12. iScience. 25, 104633 (2022).
- R. cao, Y. Zhang, SUZ12 is required for both the histone methyltransferase activity and the silencing function of the eed- eZh2 complex. Mol. Cell 15, 57–67 (2004).
- d. Pasini, A. P. Bracken, M. R. Jensen, e. lazzerini denchi, K. helin, Suz12 is essential for mouse development and for eZh2 histone methyltransferase activity. EMBO J. 23, 4061–4071 (2004).
- l. houri- Zeevi, G. teichman, h. Gingold, O. Rechavi, Stress resets ancestral heritable small RnA responses. Elife 10, e65797 (2021).
- S. varambally, S. M. dhanasekaran, M. Zhou, t. R. Barrette, c. Kumar- Sinha, M. G. Sanda,
d. Ghosh, K. J. Pienta, R. G. Sewalt, A. P. Otte, M. A. Rubin, A. M. chinnaiyan, the polycomb group protein eZh2 is involved in progression of prostate cancer. Nature 419, 624–629 (2002). - c. G. Kleer, Q. cao, S. varambally, R. Shen, i. Ota, S. A. tomlins, d. Ghosh, R. G. Sewalt,
A. P. Otte, d. F. hayes, M. S. Sabel, d. livant, S. J. Weiss, M. A. Rubin, A. M. chinnaiyan, eZh2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cells. Proc. Natl. Acad. Sci. U.S.A. 100, 11606–11611 (2003). - h. P. visser, M. J. Gunster, h. c. Kluin- nelemans, e. M. Manders, F. M. Raaphorst, c. J. Meijer, R. Willemze, A. P. Otte, the Polycomb group protein eZh2 is upregulated in proliferating, cultured human mantle cell lymphoma. Br. J. Haematol. 112, 950–958 (2001). Science AdvAnceS | ReSeARch ARticle
- t. ernst, A. J. chase, J. Score, c. e. hidalgo- curtis, c. Bryant, A. v. Jones, K. Waghorn,
K. Zoi, F. M. Ross, A. Reiter, A. hochhaus, h. G. drexler, A. duncombe, F. cervantes,
d. Oscier, J. Boultwood, F. h. Grand, n. c. cross, inactivating mutations of the
histone methyltransferase gene eZh2 in myeloid disorders. Nat. Genet. 42, 722–726 (2010). - t. J. Maures, e. l. Greer, A. G. hauswirth, A. Brunet, the h3K27 demethylase UtX- 1 regulates c. elegans lifespan in a germline- independent, insulin- dependent manner. Aging Cell 10, 980–990 (2011).
- Z. ni, A. ebata, e. Alipanahiramandi, S. S. lee, two Set domain containing genes link epigenetic changes and aging in caenorhabditis elegans. Aging Cell 11, 315–325 (2012).
- c. Jin, J. li, c. d. Green, X. Yu, X. tang, d. han, B. Xian, d. Wang, X. huang, X. cao, Z. Yan,
l. hou, J. liu, n. Shukeir, P. Khaitovich, c. d. chen, h. Zhang, t. Jenuwein, J. d. han, histone demethylase UtX- 1 regulates c. elegans life span by targeting the insulin/iGF- 1 signaling pathway. Cell Metab. 14, 161–172 (2011). - e. A. dennis, P. c. norris, eicosanoid storm in infection and inflammation.
Nat. Rev. Immunol. 15, 511–523 (2015). - P. Karlson, M. luscher, ‘Pheromones’: A new term for a class of biologically active substances. Nature 183, 55–56 (1959).
- Q. l. Wan, X. Meng, c. Wang, W. dai, Z. luo, Z. Yin, Z. Ju, X. Fu, J. Yang, Q. Ye, Z. h. Zhang,
Q. Zhou, histone h3K4me3 modification is a transgenerational epigenetic signal for lipid metabolism in caenorhabditis elegans. Nat. Commun. 13, 768 (2022). - n. liberman, M. h. Rothi, M. v. Gerashchenko, c. Zorbas, K. Boulias, F. G. MacWhinnie,
A. K. Ying, A. Flood taylor, J. Al haddad, h. Shibuya, l. Roach, A. dong, S. dellacona,
d. l. J. lafontaine, v. n. Gladyshev, e. l. Greer, 18S rRnA methyltransferases diMt1 and BUd23 drive intergenerational hormesis. Mol. Cell 83, 3268–3282.e7 (2023). - F. Y. Wang, t. t. ching, Oil red O staining for lipid content in Caenorhabditis elegans.
Bio Protoc. 11, e4124 (2021). - S. J. lord, K. B. velle, R. d. Mullins, l. K. Fritz- laylin, SuperPlots: communicating reproducibility and variability in cell biology. J. Cell Biol. 219, e202001064 (2020).
Acknowledgments:
We thank S. Strome, d. Updike, and members of the Greer lab for discussions. We thank the caenorhabditis Genetics center, which is funded by nih Office of Research infrastructure Programs (P40 Od010440) and A. Grishok for C. elegans strains.
Funding:
this work was supported by the national institutes of health grant dP2AG055947 (e.l.G.), national institutes of health grant R01AG084739 (e.l.G.), national institutes of health grant R56AG076496 (e.l.G.), charles King Fellowship (S.Y.W.), and Khorana Scholars Program (S.S.). Author contributions: conceptualization: e.l.G. and S.Y.W. Methodology: e.l.G. and S.Y.W. investigation: K.K., A.t., G.S., S.S., M.h.R., M.P.M., and S.Y.W. visualization: K.K., A.t., G.S., M.h.R., S.Y.W., and e.l.G. Supervision: S.Y.W., G.S., and e.l.G. Writing—original draft: K.K. and e.l.G. Writing—review and editing: K.K., A.t., G.S., S.S., M.h.R., S.Y.W., and e.l.G.
Competing interests:
the authors declare that they have no competing interests.
Data and materials availability:
All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. detailed analytical pipeline and all custom scripts are openly available on Github (https://github.com/kathleenkim1/Adaptation_RnA-seq_pipeline) and https://zenodo.org/record/15686193, and raw data and gene counts tables are available from the ncBi Gene expression Omnibus repository under accession number GSe255871.